 m.quanpro.cn
m.quanpro.cn爪哇犀生態(tài)和保護(hù)的新數(shù)據(jù)
爪哇犀生態(tài)和保護(hù)的新數(shù)據(jù)
New data on the ecology and conservation of the Javan rhinoceros Rhinoceros sondaicus Desmarest, 1822 (Perissodactyla, Rhinocerotidae)

一頭在烏戎庫隆國家公園內(nèi)自然棲息地的漂亮雄性爪哇犀。攝影:Tobias Nowlan
總結(jié)
盡管在原有的數(shù)據(jù)收集中投入了極大努力,爪哇犀牛(Rhinoceros sondaicus)仍是一個鮮為人知的物種。之前該分類單元遍及東南亞,從孟加拉國到越南,并且從中國南部到爪哇島。不幸的是,如今只剩大約60頭爪哇犀牛,大部分是雄性,并且全部居住在印尼爪哇島的烏戎庫隆國家公園。這種東南亞雨林里第二大的哺乳動物是低地森林里的獨居棲息者,并且不會爬上高山。它的上唇可以纏繞,并且在長度和適應(yīng)性上與吃草的印度犀(Rhinoceros unicornis)不同,爪哇犀是食性上的多面手。爪哇犀的一個突出獨特特征是頸部的“馬鞍”,而雌性的角幾乎看不見,使得性別差異即使在年輕個體中也能體現(xiàn)。基于相機陷阱數(shù)據(jù),我們推測爪哇犀也發(fā)展出了特殊和復(fù)雜的行為,以及與泥漿中打滾相關(guān)的獨特發(fā)聲。由于該物種偶爾被錯誤地鑒定為印度犀,為了在更好的區(qū)分方面提供幫助,本次工作中犀牛母子間一個獨特的行為特征被描述。也有一個目標(biāo)是強調(diào)用更多努力來了解和保護(hù)存活個體的需要,來確保這個獨特的物種不會永遠(yuǎn)消失。保護(hù)動作危險地缺乏恒定的管理控制,并且第二個種群的建立沒被當(dāng)做優(yōu)先事項考慮。作者建議增加現(xiàn)有區(qū)域的承載量,因為犀牛被限制在合適棲息地有限的烏戎庫隆國家公園內(nèi)。他們建議在無棕櫚樹的區(qū)域制造倒伏樹木來創(chuàng)造新的林中空地,以此使得幼樹生長,并增加犀牛可獲取的食物量,此外還強調(diào)了對這一珍稀物種的生存的最大化保護(hù)選項的非原位管理策略,和進(jìn)行基因銀行測序的急迫性。
簡介
這篇稿件回顧了我們對該分類單元的一些知識,包括與這個處在滅絕邊緣的物種相關(guān)的被忽略的范圍。此次分析也包含了一些關(guān)于其科學(xué)文獻(xiàn)和保護(hù)的建議。
形態(tài)學(xué)
爪哇犀體長3-3.5米,肩高1.2-1.7米,體重在1200-1500千克之間(Groves & Leslie 2011)。它的膚色通常是灰色至暗灰色,并且完全被像鱗片的特殊馬賽克狀多邊形表皮覆蓋(Lydekker 1907,Peacock 1933, Harper 1945)。它們在肢體上最明顯并且甚至在遠(yuǎn)處也能被識別。枕骨后的褶皺靠近頭部,另一處褶皺像兜帽一樣伸展穿過肩膀中部并延伸到喉部下方的一邊,幾乎組成一個連續(xù)的圓圈。另一處幾乎環(huán)繞整個身體的折疊位于肩膀后。該物種進(jìn)一步以每一條前腿上的,和一條穿過臀部的、降至大腿前方的更大橫向褶皺為特征。一個輕微的坑洼從尾根處向前延展至大腿一邊。
它的上唇是可抓握的,并且與同屬的食草者印度犀在長度和適應(yīng)性上有區(qū)別,其中爪哇犀是食性的多面手,主要取食嫩枝、幼樹和樹葉,還有很少的水果,這同樣在蘇門答臘犀牛食譜中被記錄到(Sody 1959, Groves 1982,Nardelli 2013)。該物種的食草身份完全被牙齒微磨損紋理(Hullot et al. 2019)、細(xì)觀磨損分析(Hernesniemi et al. 2011)、擴展細(xì)觀磨損方式(Taylor
et al. 2013)、半圓管定向和頭部姿勢(Schellhorn 2018)充分支持。爪哇犀可能還是以脖子上顯著的“馬鞍”最具識別性,這對該物種是決定性的(Sody 1941)。
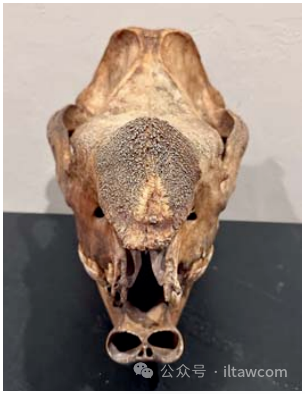


非常稀有的、帶有完整鼻骨的原版爪哇犀安南亞種(Rhinoceros sondaicus annamiticus)的頭骨,但是缺少門齒和前頜骨。這個收集于1880年的標(biāo)本被印度支那的一個傳教士交給法國的一家古董店——“R. P. Boutier的禮物”。他當(dāng)時的布道在越南河內(nèi),非常靠近中國。它在1997年被美國俄克拉荷馬市骨骼博物館獲取。攝影Jay Villemarette Sr.
盡管一些作者(Groves 1967, Cave 1969, Jan Robovsky pers. obs. in R. sondaicus inermis – Museum für Naturkunde Berlin, 2010)注意到提標(biāo)的稀疏毛發(fā)覆蓋,有一個普遍觀點是爪哇犀是無毛的。年輕個體被體毛覆蓋僅僅是個推測,毛發(fā)會隨著成熟逐漸褪去,在成體身上只留在耳緣、睫毛和尾毛(Groves 1997)。它的尾巴明顯地從后腿處突出,以至于其完整大小在側(cè)視中暴露。它們也由雌性的角識別出,爪哇犀的角僅僅是一個凸起,使得性別能在非常年輕時被確認(rèn)(Groves 1971, Groves 1982)。
我們關(guān)于該物種形態(tài)學(xué)的知識在骨骼比對方面是非常廣泛的(Groves & Leslie 2011, Asriastita 2014, Darda 2016, Schellhorn 2018, Mallet et al. 2019),但是在軟解剖當(dāng)面相當(dāng)欠缺。大腦、消化系統(tǒng)和生殖系統(tǒng)由Beddard & Treves (1887), Garrod (1877), Cave & Aumonier (1963) 和Laurie 等(1983)基于兩個樣本進(jìn)行描述,一個是Beddard & Treves(1887)檢查的來自爪哇的R. s. sondaicus亞種,另一個是Garrod(1877)檢查的源自孫德爾本斯的R. s. inermis亞種(Rookmaaker 1998)。
分布和亞種
巽他犀,或叫小獨角犀,廣為人知的名稱是爪哇犀(Rhinoceros sondaicus Anselme Ga?tan Desmarest, 1822)也被印尼語名稱提及,例如 badak或baduk Jawa, warak和Abah Gede。它的分布范圍曾經(jīng)遍及孟加拉國南部和西孟加拉邦東南部的沼澤叢林—孫德爾本斯、孟加拉中部的低地森林、印度東北部、緬甸西部和南部、泰國南部、柬埔寨、老撾、越南、馬來半島、蘇門答臘和爪哇(Loch 1937, Rookmaaker 1980, Grubb 2005),有一些跡象表明它們也曾在中國南部出現(xiàn)過(Rookmaaker 2006,Antoine 2012)。基于非常少的化石記錄,Cranbrook & Piper (2007)總結(jié)出該物種也曾在晚更新世-早全新世時期占據(jù)過婆羅洲,然而其可能的后期存活被排除(Rookmaaker 1977)。多年來爪哇犀的殘余物在印度東北部和孫德爾本斯的多個場合中被發(fā)現(xiàn)(Rookmaaker 1980, 1997,2002),所有來自不丹的犀牛報道指的是大獨角犀(Rhinoceros unicornis Linnaeus, 1758)(Rookmaaker 2016)。
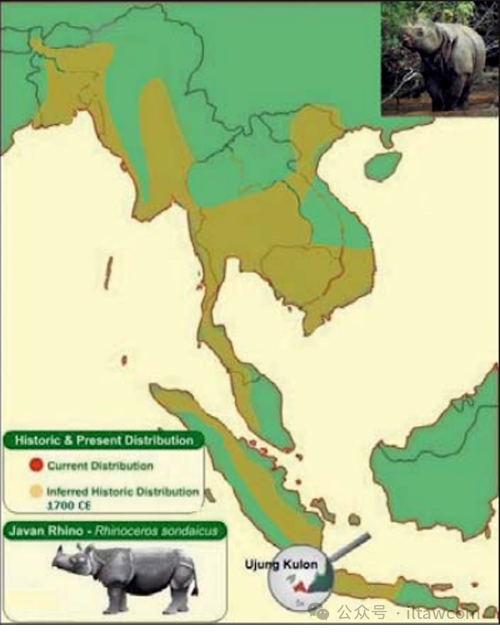
爪哇犀的分布圖。修改過的圖像來自https://badak.or.id/research-and-education/javan-rhino/
不幸的是,在三個亞種—指名亞種(R. sondaicus sondaicus)、南亞亞種(R. sondaicus inermis)和安南亞種(R. sondaicus annamiticus)—之中(Grubb 2005, Groves & Grubb 2011),只有指名亞種在爪哇島極西段的烏戎庫隆半島存活至今,那里的犀牛已達(dá)到種群密度的高峰(Nardelli 1988, 2016),而它過去曾廣泛分布于馬來西亞和蘇門答臘,以及大部分爪哇島。另一方面,南亞亞種(Lesson,1838)之前分布在孫德爾本斯和孟加拉中部、西孟加拉邦北部、印度東北部和緬甸西部,而安南亞種(Heude 1892)曾被發(fā)現(xiàn)于緬甸南部、泰國南部、柬埔寨、老撾、越南和中國南部,但是現(xiàn)在都已滅絕。
Zin-Maung-Maung-Thein等(2006)在緬甸發(fā)現(xiàn)爪哇犀的化石并且總結(jié)到該物種最有可能起源于早更新世的伊洛瓦底盆地,并且它是獨角犀屬中最原始的成員。這一發(fā)現(xiàn)表明該物種起源于亞洲大陸,并且可能在在更新世和之后的歲月中遷徙到巽他古陸。
爪哇犀的第一份系統(tǒng)地理學(xué)評估測算出指名亞種和安南亞種的分化時間大約是200萬-30萬年前(Fernando et al. 2006)。唯一的后續(xù)研究(Margaryan等2020)支持這兩個爪哇犀亞種的分化,并額外鑒定出被估計在大約54萬年前分化的北孟加拉樣本的獨特性。所有的三個被承認(rèn)的爪哇犀亞種由基因證據(jù)支持;當(dāng)務(wù)之急是獲得爪哇犀之前分布地范圍內(nèi)的細(xì)胞核或基因組數(shù)據(jù)(Moodley & Robovsky,出版中)。
考慮到爪哇犀的獨特性,值得一提的是基于整個基因組,大獨角犀和爪哇犀的分化時間被估計為430萬年前(Liu等2021)。安南亞種在越南存活至2010年(Brook et al. 2011, Brook et al. 2012)。基于腳印直徑,其頭骨尺寸被估測為指名亞種的75-80%(Polet et al. 1999, Groves 1967, Groves & Guérin 1980),表明它可能是所有三個亞種中最小的。南亞亞種可能是了解最少的,盡管可能不是最稀少的。如Rookmaaker(1997:38)記錄到:
……荷蘭、法國、英屬東印度公司的每一艘嘗試沿著恒河到達(dá)站點的船必須經(jīng)過孫德爾本斯。可以想到(而非證明)的一點是在早期人們提到印度或孟加拉的犀牛時,他們可能指的是爪哇犀而不是“大獨角犀(Rhinoceros unicornis)”……
根據(jù)Spartaco Gippoliti (私人通信2023),爪哇犀在過去可能經(jīng)常被錯誤鑒定為大獨角犀。海運以及蘇伊士運河在1869年的開通促進(jìn)了長途貨運,使得動物園和博物館更容易獲取遠(yuǎn)處的標(biāo)本。

越南吉仙國家公園最后的爪哇犀安南亞種之一。盡管2009-2010年間野外調(diào)查(Brook等2012)沒有總結(jié)性地指出爪哇犀在越南的絕跡,調(diào)查的結(jié)果強烈表明這一分類單元的滅絕。攝影:WWF越南
因此在19世紀(jì)晚期和20世紀(jì)早期,隨著來自印度和孟加拉的船駛向的里雅斯特(當(dāng)時屬于奧匈帝國)的港口,他們也會給動物經(jīng)銷商帶去額外的貨物,像是在那個港口有一個專業(yè)的活體動物倉庫的Carl Hagenbeck (Hagenbeck 1912: 148)。在那時,從靠近孟加拉灣附近的孫德爾本斯采集犀牛可能是更實際的,這是由于很少有大獨角犀能被活著帶到印度的港口,因為這一物種只在印度次大陸北部出現(xiàn)。這一假設(shè)被一個事實確認(rèn),那就是那不勒斯(意大利)的一頭確定在那里死亡的被推定為大獨角犀,而大學(xué)動物學(xué)博物館最近重新認(rèn)定為爪哇犀(Improta & De Francesco 2022)。博物館獨角犀屬樣本的系統(tǒng)普查肯定是值得的(Robovsky& Rookmaaker 2022)。

那不勒斯(意大利)腓特烈二世大學(xué)動物學(xué)博物館里的爪哇犀骨架。犀角被盜但是以一個過長的復(fù)制品替代。攝影:那不勒斯動物學(xué)博物館
行為
考慮到爪哇犀的稀有性以及沒有圈養(yǎng)下的繁殖(Rookmaaker 1998),目前關(guān)于該物種繁殖生態(tài)和社會行為的知識是有限的。這個問題又進(jìn)一步因為居住在受控環(huán)境下活體的缺乏而加劇,使得直接的觀察具有挑戰(zhàn)性。由于爪哇犀神秘的天性和害羞,對其繁殖行為的了解非常少(Groves & Leslie 2011)并且目前的數(shù)據(jù)大部分以關(guān)于少數(shù)個體的早期說明為基礎(chǔ)(Hoogerwerf 1970, Ammann 1986)。根據(jù)Gokkon(2020),仍有74頭動物存活,并且全部孤立地生活;研究該物種是困難的(Dinerstein 2011)。在2019年,基于相機陷阱識別,烏戎庫隆國家公園的爪哇犀種群包括28頭成年雄性(41.2%)、23頭成年雌性(33.8%)、8頭年輕雄性(11.8%)和9頭亞成年雌性(13.2%)(Putra等2020)。根據(jù)Putro(2023),76頭犀牛外加3頭幼崽在2022年出現(xiàn)。一項近期報道正在挑戰(zhàn)這些數(shù)據(jù)(見下文的保護(hù)一節(jié))。
根據(jù)一些記錄,爪哇犀在18世紀(jì)時在該國普遍存在(Groves 1967),并且對它的狩獵得到爪哇政府支持(Ramono等1993)。然而,即使在唐朝時期(公元618-906年),犀角出口是爪哇政府的一項重要收入來源,這可能促進(jìn)了該物種的消亡(Nardelli 1988)。
Nardelli(2016)近期發(fā)表意見稱爪哇犀并不是一個合群的物種,并且該物種不是山地居民,數(shù)個研究者做出這樣的宣稱(Horsfield 1824, Sody 1941,Hoogerwerf 1970, Ellis & Talukdar 2020, Britnell et al. 2021)。
如之前所記錄,考慮的雌性更具社交性,它們的領(lǐng)地重疊相當(dāng)多,但雄性傾向于阻擋其他雄性來保護(hù)它們的領(lǐng)地并可能只在邊緣有重疊。這一行為像蘇門答臘犀牛的行為,但與半開放草原的大型食草占用者的大獨角犀中記錄到的相反。一個典型的雄性領(lǐng)地范圍從12到21平方公里,而雌性的僅僅覆蓋4至13平方公里(Ammann 1986)。
因此,我們今天所知道的關(guān)于該物種的大部分信息來源于它們被研究的痕跡(例如腳印、尿液噴射和糞便),這只能揭示其實際繁殖生態(tài)的很少內(nèi)容(Horsfield 1824, Sody 1941, Hoogerwerf 1970, Ammann 1986)。盡管如此,Ammann(1986)對幾頭母犀牛及其幼崽的觀察表明它們組成永久的聯(lián)系。作者進(jìn)一步推測雌性幼崽會和母親留在一起,而雌雄犀牛間的聯(lián)系會在4-5天后中斷,這可能是與雌性動情期持續(xù)時間對應(yīng)的時間跨度。
這些發(fā)現(xiàn)近期被Wilson(2021)證實,在初次的一些觀察里,他證實雄性和雌性爪哇犀都更偏好獨處。根據(jù)這個作者廣泛的論文,社交行為大部分局限在打滾的地點,它們充當(dāng)用于體溫調(diào)節(jié)目的,以及為與其他犀牛交流、見面和互動提供機會的共享棲息地。因此,爪哇犀傾向于在打滾地中和邊上發(fā)出相比在森林棲息地里頻繁3倍的聲音(并使用不同的呼叫聲音)。盡管帶崽的雌性爪哇犀被發(fā)現(xiàn)與其他雌性犀牛及其幼崽建立社交紐帶,這樣的社交互動通常不會擴展至成年雄性,除非雌性在性行為上是樂于接受的。出于這個原因,雄性爪哇犀有規(guī)律地拜訪打滾地。根據(jù)Wilson(2021),雌性在靠近打滾地和林中空地時明顯更警覺,并且甚至在雄性早已出現(xiàn)時轉(zhuǎn)頭逃跑。

爪哇犀的打滾泥坑。攝影:國際犀牛基金會
Wilson(2021)對打滾地中和附近拍攝的相機陷阱視頻中的犀牛語音呼叫的分析揭示出7種發(fā)聲描述符號還有附帶的聲譜圖。Wilson(2021)也能夠確認(rèn)所有性別在飲水,或者品嘗或攝入泥土后后展示出裂唇嗅反應(yīng),作者將其歸因于確認(rèn)性別,以及打滾地使用者的繁殖和主導(dǎo)狀況的本能需要。因為被犀牛頻繁光顧的打滾地通常被植被很好地隱藏,通常形成為雨水能貯存的6-7米×3-5米的尺寸,犀牛媽媽在帶著幼崽進(jìn)入前能在相對安全的位置監(jiān)視它們。
爪哇犀幼崽傾向于跟在媽媽身后(Nardelli 2017)。然而,這一行為也在蘇門犀和黑犀牛幼崽中被觀察到(Joubert & Eloff 1971; Benda et al. 2020, Van Strien 1986)。另一方面,白犀牛和大獨角犀幼崽走在媽媽前面(Owen-Smith 1973, 1975, Laurie 1978, 1982),像Hitchins(1986)解釋的那樣,這使得它們更不可能成為獅子(Panthera leo)、斑鬣狗(Crocuta crocuta),以及老虎(Panthera tigris)的受害者(Dinerstein 2003)。這些觀察應(yīng)該導(dǎo)向食葉犀牛的幼崽跟在媽媽身后(使得媽媽清理粗糙的植被并暴露出可食得的葉和芽),而食草犀牛的后代走在前面(使幼崽在開闊地帶捕食者的實現(xiàn)中變模糊)的結(jié)論。

有幼崽跟隨的雌性爪哇犀,在此網(wǎng)頁的視頻中更明顯https://www.facebook.com/watch/?v=529657705848991
進(jìn)一步的保護(hù)由母親和幼崽間幾乎聽不見的發(fā)聲所提供,因為該物種的幼崽偶爾被爪哇豹(Panthera pardus melas)或群居的爪哇豺(Cuon alpinus sumatrensis)所捕食(Wilson 2021)。據(jù)信爪哇虎(Panthera tigris sondaica)之前可能也會捕食年輕的爪哇犀,并且可能造成其種群相當(dāng)大的減少。
由于爪哇犀要求相當(dāng)多的食物,種群密度應(yīng)該被減少到更有生產(chǎn)力的程度(Wilson 2021)。然而這實現(xiàn)起來是困難的。雌性大獨角犀首次繁殖的年齡估計是7-7.5歲,表明對爪哇犀來說可能是相同的(Dinerstein 2003)。由于其小種群,這意味著只有部分雌性爪哇犀是同時在繁殖上活躍的。另一方面,它的長壽可能對種群恢復(fù)有幫助。盡管如此,這些推測只能通過進(jìn)一步研究證明或駁斥(Wilson 2021)。
相似地,目前不知道爪哇犀是像大獨角犀、黑犀和白犀那樣的自發(fā)性排卵,還是像蘇門犀那樣由交配引起(刺激)的排卵(Roth et al. 2001)。
妊娠期同樣未知,并且目前被預(yù)計為16個月,考慮到交配很少被觀察到,這使得受孕的計時很困難。Dinerstein(2023)認(rèn)為大獨角犀在野外每2.5年可以繁殖一個幼崽。
當(dāng)準(zhǔn)備好交配時,雄性和雌性參加能簡易識別的求愛行為,由此雄性發(fā)出重復(fù)的“短褲(short-pant)”呼叫,雌性對此回應(yīng)發(fā)出重復(fù)的“moo-bray”呼叫。在靠近時,雄性會用它們的角摩擦雌性頸部下方。如果樂于接受的話,雌性會通過張嘴和用背部摩擦雄性做出回應(yīng)(Wilson 2021)。根據(jù)(Wilson 2021),雄性和雌性都會和幾個伴侶交配(一夫多妻)。
如之前討論所示,爪哇犀和大獨角犀的對比可能會導(dǎo)向錯誤的結(jié)論,因為兩個分類單元有不同的生態(tài)。因此,當(dāng)制定保護(hù)策略時,注意到爪哇犀需要廣袤的、連續(xù)的低地雨林用以重新住入是極其重要的,并且這樣的棲息地目前只在蘇門答臘的少數(shù)地方存在。作為巨型動物群最后幸存者之一的亞洲犀牛對于生態(tài)恢復(fù)也十分重要(Louys等2014)。
食性、棲息地和生態(tài)角色
爪哇犀是東南亞雨林中第二大的哺乳動物并且高度依賴低地森林,因為森林的密度保護(hù)這些敏感的動物免于陽光,并且確保穩(wěn)定的水源供應(yīng),而樹木提供足夠的幼樹和葉片,是其主要的食物來源。該物種是一種全才型食葉者(僅僅偶爾被發(fā)現(xiàn)于農(nóng)田周邊),但是不像曾經(jīng)重疊分布的蘇門犀(Dicerorhinus sumatrensis,F(xiàn)ischer 1814)—同樣是食葉者和雨林居民—裝配有“緊握形狀蹄子”,爪哇犀無法在山坡爬上爬下。
爪哇犀主要取食嫩枝和樹葉,每天攝取大約80-100千克的食物(例如蘇門犀吃60-70千克,體重700-900千克),這對于單只動物去獲得是具有挑戰(zhàn)性的(個人觀察)。
爪哇犀目前占據(jù)爪哇的低地常綠次生林,并且它曾在越南占據(jù)邊緣棲息地(Groves & Leslie 2011)。但是考慮到其歷史和史前分布(Groves 1967, Groves & Leslie 2011),它有可能占據(jù)與地域典型的植被和低地/山地氣候相關(guān)的更多樣棲息地(Morley 2018)。將東南亞植被地圖(Morley 2018)和爪哇犀地點結(jié)合(Groves 1967),它曾占據(jù)過雨林、季節(jié)性常綠林、半常綠林和濕潤落葉林。
爪哇犀似乎符合難民物種的標(biāo)準(zhǔn)(Kerley等2012),像是:在過去分布更廣,目前以非常低的數(shù)量出現(xiàn)在非常有限的地理范圍,并且居住在一定程度上質(zhì)量不理想的一個位置(Griffiths 1993)。
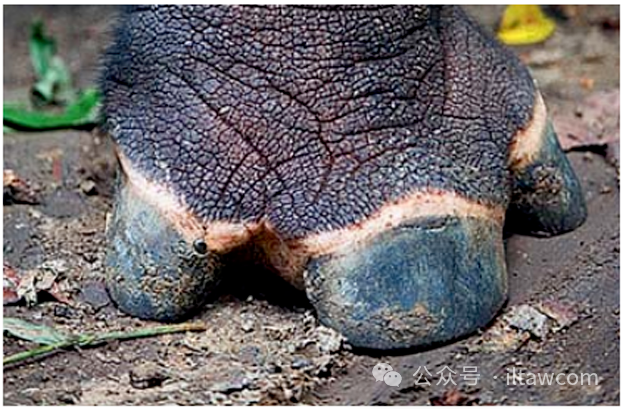

犀牛足部對比—第一幅:蘇門犀;第二幅:爪哇犀—一個獨特安裝的部位。
第一幅攝影:Mark Carwardine/naturepl.com
第二幅攝影:David Ayre
保護(hù)
目前,爪哇犀個體的數(shù)量停滯不前—如果沒有減少—可能是由于不足的棲息地和種群管理措施、盜獵和近親繁殖,以及繁殖期雌性極低數(shù)量導(dǎo)致的有限繁殖力。特別是隨著人口持續(xù)增長,設(shè)法解決這些問題的失敗會進(jìn)一步削弱現(xiàn)有管理實踐的價值,由此將早已破碎的犀牛種群置于更大的滅絕風(fēng)險。因此,現(xiàn)存的保護(hù)和保護(hù)策略需要被修改,其中主要重點是阻止盜獵,這被犀角在亞洲的高需求持續(xù)驅(qū)動。
為了克服這些挑戰(zhàn),現(xiàn)有保護(hù)區(qū)的承載量也需要增加。例如,爪哇犀被局限在120000公頃的烏戎庫隆國家公園內(nèi)范圍內(nèi),但根據(jù)Griffiths(1993),只有被分成三塊主要飛地的30000公頃的園區(qū)被認(rèn)為適合犀牛(Manurung等2023)。此外,這些棲息地里的60%(18000公頃)出于最佳水平,剩下的40%(12000公頃)是次優(yōu)質(zhì)量(Griffiths 1993),主要由于入侵的鈍葉羽棕(Arenga obtusifolia)形成的棕櫚林、省藤屬(Calamus)和露兜樹屬(Pandanus)。考慮到這一評價包括沒有發(fā)現(xiàn)犀牛的Gunung Payung地區(qū),很明顯從長遠(yuǎn)來看,沒有改進(jìn),爪哇犀在烏戎庫隆國家公園內(nèi)的存在就無法被保證。
某些補充飼料能否幫助提高這一爪哇犀最后種群的健康、存活和繁殖是個問題—即新的林中空地能被幾棵倒下的大樹創(chuàng)造,在相距甚遠(yuǎn)的棕櫚林地,這樣的林中空地能生長出來以增加犀牛可獲取的食物量。倒樹留在樹蔭中的空間允許更多陽光到達(dá)地標(biāo)并創(chuàng)造出其他植被生根和繁茂的空間。如果一棵樹帶著其他植物倒下,這更加能夠幫助周邊的重新生長。這個新計劃能以非常低的代價被立刻實施。

帶有兩年重新生長幼樹的倒樹。攝影:Amphoe Muang Phatthalung

取食倒樹附近幼樹的雌性爪哇犀。鈍葉羽棕(Arenga obtusifolia)現(xiàn)在占據(jù)公園內(nèi)許多地方的雨林,并且通過限制犀牛取食植物的生長減少了犀牛的采食。攝影:印尼烏戎庫隆國家公園和林業(yè)環(huán)境部
更深層次的問題來自從不足的保護(hù)到盜獵,因為犀牛保護(hù)單元和受物理安全控制委托的公園保安(Haryono等2015)沒有力量巡邏如此大的區(qū)域。相機陷阱被安裝在公園各處用以觀察和統(tǒng)計犀牛,這些明顯只能鑒定安全漏洞,而非阻止未授權(quán)的訪問。不過,由于今年猛增的專業(yè)盜獵的程度,任何出版的官方報道數(shù)據(jù)能很快改變。有證據(jù)表明這些罪犯已經(jīng)從蘇門答臘轉(zhuǎn)移到烏戎庫隆國家公園(Manurung等2023)。例如,一頭雄性犀牛在2018年被殺,并且當(dāng)尸體被追回時,一個彈孔在頭骨上被發(fā)現(xiàn),而在某個不明確的年份,相機拍到一頭背部有兩個孔洞的受傷犀牛。
因此,所有利益相關(guān)者的堅定承諾是確保烏戎庫隆國家公園能以被估計為50-60頭個體的穩(wěn)定最大承載量運行的關(guān)鍵保證(Ammann 1986, Nardelli 1986, Santiapillai 1990, Fernando et al. 2006, Haryono et al. 2016)。盡管這一數(shù)據(jù)在1980年代被建立,它與Manurung等(2023)做出的最近期的評估相一致。
指向利益相關(guān)者行動的急迫性的深層證據(jù)能在一個當(dāng)?shù)胤钦M織— Auriga Nusantara—在2023年4月出版的報道中被發(fā)現(xiàn),表明存活的爪哇犀數(shù)量被印尼政府機構(gòu)嚴(yán)重高估(Manurung等2023),至少有15頭在近期沒有被遍及公園的220臺相機拍到。
因此,目前只有60頭個體占據(jù)烏戎庫隆國家公園(Manurung等2023),并且考慮到只有23頭是雌性(Haryono等2015),有一種可能是如今后者當(dāng)中只有大約12頭能成功繁殖,種群無力增長。
盡管對抗任何盜獵活動仍然是最緊迫的問題,爪哇犀的基因多樣性也處在一個危險狀態(tài)。Fernando & Melnick(2003)和Fernando等(2006)進(jìn)行的基因研究表明爪哇犀種群接近純合,只有兩條等位基因變異和兩個母系支系(單倍型),由此只有全部種群的14%攜帶2號單倍型。近期對爪哇犀形態(tài)學(xué)特征的觀察也表明13頭個體受近交衰退導(dǎo)致的出生缺陷的影響(Putro 2023).
因此,未來旨在烏戎庫隆中的爪哇犀保護(hù)的努力必須聚焦在保護(hù)它們的安全、提高棲息地質(zhì)量和提高基因活力方面,因為這些問題一旦解決失敗將會導(dǎo)致種群停滯或減少。盡管一些研究者和利益相關(guān)者都認(rèn)為讓所有的爪哇犀種群局限在同一個半島可以促進(jìn)更大程度的保護(hù),但棲息地多樣性的缺失也增加了環(huán)境危機和疾病爆發(fā)的風(fēng)險。因此,為了限制任何基因變異的影響(Stephens & Sutherland 1999)和阻止快速地疾病傳播,爪哇犀應(yīng)當(dāng)被提供可替代的地點(Wilson 2021)。
如一些作者所建議(Ammann 1986, Nardelli 1986, 2016, Santiapillai & Suprahman 1986,Seal & Foose 1989, Haryono et al. 2016),一個受監(jiān)管的種群應(yīng)當(dāng)在一個可替代的地點被建立。盡管Leader-Williams (1993)告誡說圈養(yǎng)犀牛的種群增長率傾向于比良好保護(hù)的野生種群更低,這是目前唯一可行的觀點。一個作為更廣泛的保護(hù)倡議的一部分的非原位繁育項目會極大貢獻(xiàn)爪哇犀的恢復(fù),并且一旦取得足夠的進(jìn)展,其他觀點可能會被考慮,比如在最佳棲息地內(nèi)成對引入。具體來說,這樣的舉措已經(jīng)成功用于大獨角犀,導(dǎo)致了該物種異質(zhì)種群的顯著增長,這是繁殖率提高和不同地點新種群建立的結(jié)果。盡管如此,因為獨角犀屬的生態(tài)與雙角犀屬不類似,無法理所當(dāng)然地認(rèn)為能有相同的成就。
當(dāng)考慮上文概述的替代地點時, Auriga Nusantara報道(Manurung等2023: 28)提供的建議應(yīng)當(dāng)作為寶貴的指導(dǎo):
1.爪哇犀和烏戎庫隆國家公園的保護(hù)方面應(yīng)該有整體改進(jìn);
2.國家公園辦公室和/或環(huán)境林業(yè)部應(yīng)該按照學(xué)術(shù)標(biāo)準(zhǔn)(并使用多種并行方法)計算出爪哇犀種群;
3.對烏戎庫隆國家公園辦公室進(jìn)行一次全面評估,檢查埼機構(gòu)、預(yù)算和項目方面的安排;
4.認(rèn)真地實施一個爪哇犀第二種群或第二棲息地的項目;
5.鼓勵和開放對爪哇犀研究的空間。
實施上述措施時,必須找到擴展爪哇犀種群極限的有效方式,以此防止未來不可避免的基因崩潰。另外,如果在爪哇找不到足夠大的、能為更多數(shù)量的爪哇犀充當(dāng)優(yōu)質(zhì)棲息地的合適區(qū)域,蘇門答臘應(yīng)當(dāng)被考慮為替補。當(dāng)為安置爪哇犀選擇新地點時,要考慮的主要因素是免受疾病和盜獵的安全性,和至少50頭的承載量。這被視為確保種群增長的最低個體數(shù)(Haryono等2016)。
如概述所說,急需一個緊急的非原位行動計劃來拯救爪哇犀免于滅絕。這只能通過所有利益相關(guān)者的真正關(guān)注和承諾來實現(xiàn),尤其是那些有政策決定力的人—和公正的意見—因為他們的支持對于提高現(xiàn)有的管理能力至關(guān)重要。然而,目前的關(guān)注點似乎聚焦在將所有爪哇犀保留在爪哇島,并且解決與盜獵相關(guān)的最直接的擔(dān)憂。這一短視的觀點未能解釋同一區(qū)域內(nèi)近親繁殖導(dǎo)致的基因多樣性缺失帶來的風(fēng)險,這只能通過建立一個第二種群來克服,至少是如此。因此,國家政府的強力領(lǐng)導(dǎo)對于為這一高度瀕危的物種提供存活,和在未來潛在地繁榮的機會,是極其重要的。
如Maguire等(1987)指出,非原位繁育給瀕危物種在野外提供足夠棲息地不總是最佳替代品。盡管如此,保障繁殖群體的存在是基本的。在爪哇犀的案例中,大約60頭個體居住在一個沒有任何自然擴散機會的小的飛地中,一個非原位的策略是最安全的可行選項(Nardelli 1986, Seal & Foose 1989)。而每個措施應(yīng)該被采取以維持烏戎庫隆國家公園內(nèi)的爪哇犀處于最高密度,上述的危險將仍然存在,除非一個次級地點被建立(Wilson 2021)。
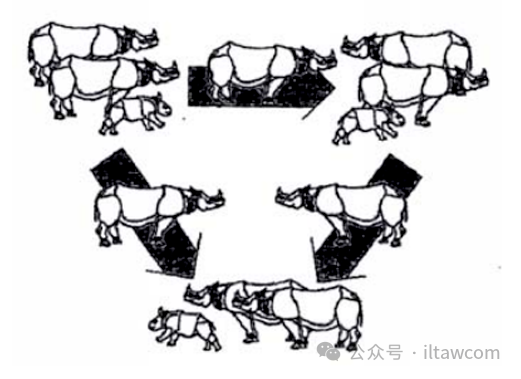
用以維持異質(zhì)種群間基因流動的亞種群間受管控的遷移。制圖:Tom Foose
這些論點由近期的Auriga Nusantara研究(Manurung 等2023, Gokkon 2023)所支持,其發(fā)現(xiàn)證實維持維持現(xiàn)狀只會適得其反。進(jìn)一步證實其有效性的證據(jù)能在馬來西亞沙巴州和印度尼西亞被找到,在那里幾個蘇門犀種群在過去30年間滅絕。情況正在急轉(zhuǎn)直下,而且更先進(jìn)的戶外技術(shù)證明野外種群正處在危急關(guān)頭,并需要大的干預(yù)。

取食樹葉的蘇門犀。攝影:Alain Compost
相反地,在一處單一設(shè)施內(nèi)的蘇門犀非原位種群正繁榮到相當(dāng)?shù)某潭龋灾劣谠谝粋€不同于原地區(qū)的安全位置建立蘇門犀保護(hù)區(qū)變得很有必要。非原位管理計劃的終極目的是投放到周邊森林的非原位繁育的犀牛產(chǎn)生出一個野生種群。然而,說到這里,沒有用于未來放生的地點被確定下來,因為目標(biāo)也有避免在蘇門答臘與蘇門犀、大象競爭資源,或者被老虎捕食。
同樣值得記錄的是人工繁育技術(shù),例如人工授精,已經(jīng)足夠先進(jìn),但它目前只在管理良好的設(shè)施上可行。盡管如此,如果爪哇犀種群因為疾病、衰老或其他因素迅速減少,需要干細(xì)胞和生物技術(shù)的人工繁育技術(shù)是非常重要的。根據(jù)Hildebrandt等(2021),人工授精早已在超過50個原生物種中制造出可生長發(fā)育的后代,包括兩頭犀牛。細(xì)胞培養(yǎng)的生物銀行是非常有前景的保護(hù)選項,因此對它們的保存應(yīng)當(dāng)被高度推薦,就像在蘇門犀的案例中那樣(Brandt等2018)。

爪哇犀頭部特寫。在顯眼的皮膚上能清晰看見,完全被像魚鱗的特殊表皮馬賽克狀多邊形覆蓋,還有可纏繞的上唇。犀角前端擁有整個屬中典型的溝槽(Groves 1971)。攝影:Tobias Nowlan
成問題的性別比
研究的大體量表明滅絕的風(fēng)險在種群低于特定門檻時激增,因為阿利效應(yīng)(Stephens & Sutherland 1999)和隨機性(Courchamp等1999)。另外,隨著種群大小下降,基因池也隨之下降,會因為隨機漂變和近親繁殖,而增加遺傳缺陷和有害突變累積的傳播可能(Seal & Foose 1989, Allendorf & Luikard 2007)。
而個體的健康水平能通過操縱性別比提高,這樣的干預(yù)需要僅能在非原位實現(xiàn)的專門設(shè)施。說到這里,在其他分類單元中,通過諸如那些旨在調(diào)整性別決定生態(tài)或社會因素的非入侵式方法,已經(jīng)取得一些成功。盡管如此,侵入式方法—包括胚胎激素治療或在輔助生殖技術(shù)前確定精子性別—在實踐中更流行,因為它們傾向于產(chǎn)出更可靠的結(jié)果(Wedekind 2012)。
根據(jù)Haryono等(2015),有著35頭雄性和23頭雌性,爪哇犀種群的雄/雌比例在2012年為1:0.76,降至2013年的1:0.66。對更稀少的雌性的相同觀察已經(jīng)由Hoogerwerf (1970)得出。在孤立種群中,性別比偏差和滅絕風(fēng)險是非常重要的保護(hù)問題(Clout et al. 2002, Grayson et al. 2014, Barrientos 2015),并且在爪哇犀的案例中應(yīng)該被嚴(yán)肅評估。
然而,由于更近的數(shù)據(jù)無法獲取,很難外推這個下降趨勢以形成可靠的推斷。這個問題進(jìn)一步因一個事實惡化,那就是盡管有建議,雌性爪哇犀被不斷地和在個體層面被監(jiān)控(Nardelli 2016),即便有220臺陷阱相機被部署在隨機地點來確定總數(shù)—這個數(shù)據(jù)對外行來說是發(fā)自內(nèi)心的。
由于雌性的數(shù)量決定任何物種的潛在種群增長,擁有過多的雄性爪哇犀是一個主要的種群統(tǒng)計問題。它也增加了對配偶的競爭和對繁殖搭檔的挫敗,導(dǎo)致嚴(yán)重受傷和心理從屬方面的有害結(jié)果的性沖突。
因此,為了這一接近滅絕物種的繁殖率增加,性別比操縱可能被需要。同樣值得記錄的是在原位的野生非洲犀牛能夠適應(yīng)后代的性別,因此自然地限制了繁殖競爭并維持健康的性別平衡(Berkeley 2011)。這些初步的發(fā)現(xiàn)可能表明相似的性別控制機制對爪哇犀來說是可能的,假如它們在被給定非原位設(shè)施的情況下,在那里這些補償性的回應(yīng)能夠開始。總之,因為雄性繼續(xù)在爪哇犀原位種群中占主導(dǎo),基因惡化時不可避免的,證明了對這一最珍貴的物種生存的非原位管理策略的急迫性。
另一個問題是圈養(yǎng)犀牛的繁殖生態(tài)是繁殖條件的優(yōu)化,像是雄性的轉(zhuǎn)移、繁殖群體的大小等等(Scott等2023)。在該物種潛在的圈養(yǎng)繁殖方面,基于該物種已知的生態(tài),和其他犀牛物種在圈養(yǎng)條件下的經(jīng)驗,雄性的交換和分開應(yīng)該被準(zhǔn)確評估。
理解爪哇犀生態(tài)的觀點
總結(jié)我們的回顧,我們推薦這些主題來記錄爪哇犀的生態(tài)參數(shù):
1.在形態(tài)和基因研究方面,爪哇犀的保存標(biāo)本和基因銀行樣本的目錄應(yīng)當(dāng)被推薦;目前爪哇犀保存標(biāo)本唯一可獲取的列表在1930年代出版(Barbour and Allen 1932, Loch 1937);
2.因為爪哇犀的解剖學(xué)參數(shù)受樣本數(shù)量限制,并且僅限于器官系統(tǒng),在未來測量所有可獲取標(biāo)本是值得的,因為它們在死亡個體上保存盡可能多的身體部分。研究爪哇犀的形態(tài)和解剖學(xué)特征提供對其身體適應(yīng)性的了解,例如它獨特的皮膚褶皺。該物種中可能雌性更大的兩性異型(Groves 1982回顧)是一個需要基于更大樣本數(shù)量驗證的問題。其骨骼系統(tǒng)、繁殖器官和其他解剖特征的細(xì)致檢驗有助于理解它的生理能力和繁殖生態(tài);
3.對每個可獲取的爪哇犀活體細(xì)胞培養(yǎng)的生物銀行測序有極大保護(hù)價值,如Brandt等(2018)在蘇門犀的案例中,Saragusty等(2016)和Korody等(2021)在北部白犀牛的案例中所解釋的那樣;
4.因為爪哇犀是最后一種沒有已知核型的犀牛(Houck et al. 1994, Trifonov et al. 2012),細(xì)胞培養(yǎng)可用于染色體參數(shù)的細(xì)致檢測;
5.盡管在爪哇犀發(fā)聲項目的文獻(xiàn)中有一些當(dāng)前的進(jìn)展(Wilson 2021),次聲未被記錄;需要澄清的是爪哇犀是否是唯一沒有次聲的犀牛物種(von Muggenthaler et al. 1993);
6.因為同位素證據(jù)可以證實一些食性和棲息地轉(zhuǎn)變,還有一些物種的難民物種特征(Kaczensky et al. 2017, Suraprasit et al. 2020; Isarankura Na Ayudhyaet al. 2022),來自歷史和現(xiàn)有種群的同位素證據(jù)對于理解該物種的原有棲息地和食性偏好極其重要;
7.基因研究在理解爪哇犀的種群動態(tài)、基因多樣性和與其他犀牛物種的關(guān)聯(lián)性中起到關(guān)鍵作用。分析更多的爪哇犀基因組能提供關(guān)于它的進(jìn)化史、潛在基因脆弱性和導(dǎo)致其減少的因素的寶貴信息。
8.調(diào)查爪哇犀的行為和生態(tài)有助于理解其棲息地需求、覓食習(xí)慣、社會結(jié)構(gòu)和交配行為。研究它的移動、活動模式和偏好的棲息地類型能幫助鑒定出重要保護(hù)區(qū)域和設(shè)計有效的管理策略;
9.爪哇犀的繁殖生態(tài)在保護(hù)中是最重要的。對婚配制度、繁殖成功、妊娠期和影響繁殖的因素的研究能為提高該物種的繁殖成功率指明努力方向,對圈養(yǎng)和野外都如此;
10.監(jiān)控爪哇犀的健康狀況對于鑒定和緩解潛在威脅是重要的,并入疾病或寄生蟲。研究它們的免疫系統(tǒng)、對特定病原體的敏感性,以及與棲息地退化和與人類互動相關(guān)的健康風(fēng)險能夠影響保護(hù)策略,包括疾病預(yù)防和管理;
11.理解爪哇犀的棲息地需求,包括偏好的植被類型、水資源和景觀連接性,對有效的管理計劃是重要的。研究棲息地喪失、破碎化和退化對該物種的影響能為棲息地恢復(fù)指明努力方向,并影響保護(hù)區(qū)管理的策略。

雄性爪哇犀在最喜歡的要素中:水。在這幅圖里,可以觀察到“馬鞍”特征,幾乎完整的多邊形表皮生長覆蓋,以及角前端的溝槽。攝影:Tobias Nowlan
結(jié)論
因此,Haryono等(2016)建議用關(guān)系相對遠(yuǎn)的4頭雄性和4頭雌性開始爪哇犀的原位項目,這篇論文的作者認(rèn)為這個組合對非原位項目也是充足和有活力的繁殖集群。作者證實盡管這個計劃進(jìn)一步推薦優(yōu)先建立2個非原位設(shè)施,兩對犀牛分到每一個當(dāng)中,因為非原位繁殖是建立第二個爪哇犀種群最安全和最有成果的選項,正如第一個蘇門犀項目和保護(hù)區(qū)經(jīng)歷的那樣。出于這個目的,適合繁殖的動物應(yīng)該被選擇性鑒定,在這之后的,已在蘇門犀身上成功實施的捕捉、適應(yīng)環(huán)境的和管控可以被采用。不過,在抓捕期間,必須采取關(guān)照來避免其他個體的過度干擾并避免抓帶崽雌性。由于爪哇犀居住在烏戎庫隆國家公園內(nèi)相對接近的地方,每年應(yīng)該抓捕一頭犀牛。免去了干擾,這種交錯式抓捕也考慮到在擴大繁育項目前任何協(xié)議的細(xì)化。
具體來說,任何非原位地點的生態(tài)、行為挑戰(zhàn),或合并群體與繁殖犀牛的問題,以及抓捕和轉(zhuǎn)移的物流,應(yīng)該被定期檢查,以此用及時的方式解決任何挑戰(zhàn)。一旦兩個地點被建立,動物的戰(zhàn)略性交換也被建議,因為這會極大提高基因多樣性。
在這些倡議同時,烏戎庫隆國家公園的保護(hù)手段需要現(xiàn)代化和強化,來預(yù)防盜獵和疾病傳播。考慮到盜獵組織增加的復(fù)雜度,以及當(dāng)?shù)丶倚罂拷珗@內(nèi)野生動物的地區(qū),保持現(xiàn)狀能將犀牛置于無法挽回的種群損失的風(fēng)險(Manurung等2023)。
烏戎庫隆國家公園
爪哇犀是不幸的“依賴保育”物種(Goble et al. 2012, Scott et al. 2010)。當(dāng)一個物種面對的威脅無法被消除,只能被管理時,該物種便是依賴保育。有兩種形式的保護(hù)依賴:種群和威脅管理依賴—其生存取決于旨在管理它面對的威脅的保護(hù),考慮到壓力無法被完全消除,而只能管理的這樣一個增加的共識。因此,任何旨在阻止這一高度瀕危物種滅絕的倡議必須以謹(jǐn)慎的保護(hù)計劃開始,這應(yīng)該包括對依賴保育所在處,或被設(shè)計為最清晰的地點的鑒定。
這些計劃也應(yīng)該并入存活種群的維持,即使生態(tài)恢復(fù)目標(biāo)達(dá)成,持續(xù)的物種特異性干預(yù)是需要的。這些論據(jù)適用于所有“保育依賴”物種,不單單是爪哇犀。盡管如此,對于存活在野外的爪哇犀,每個個體在其整個生命周期內(nèi)需要完全和專職的人工監(jiān)督。另外,爪哇犀早已是“難民物種”(Kerley等2020)。難民物種被定義為無法獲取最佳棲息地,被限制于次優(yōu)棲息地內(nèi),后果是減少的健康和密度,和隨之而來的保護(hù)風(fēng)險。在這個和其他自然棲息地消失或者被人不利影響的物種一樣的案例中,爪哇犀個體生活的地區(qū)不僅在大小和資源多樣性上受限,也對它們的活動、食性和繁殖造成限制。所有這些不利影響已經(jīng)導(dǎo)致該物種的健康和密度與歷史記錄相比的顯著遞減(Kerley等2012)。同樣值得記錄的是實施在次優(yōu)棲息地內(nèi)爪哇犀的保護(hù)努力傾向于收益有限,因為其提高健康和密度的能力在這種情況下嚴(yán)重受限(Kerley等2020)。在這種環(huán)境下,“基線移位綜合癥”(Pauly 1995)—基線移位綜合癥描述因為缺乏人類經(jīng)驗、記憶和/或其過去情況的知識的自然環(huán)境的情況的公認(rèn)標(biāo)準(zhǔn)的逐步改變—也起到了作用,因為在評估系統(tǒng)狀態(tài)和其在特定時期內(nèi)經(jīng)歷的改變時,未能采取客觀措施可能會導(dǎo)致一個保護(hù)管理活動是實際有效的錯誤結(jié)論錯誤的結(jié)論,甚至難民物種被限制在邊緣棲息地(Kerley等2012)。根據(jù)Caughley (1994: 229),對于經(jīng)歷范圍收縮的受威脅物種而言這種風(fēng)險尤其嚴(yán)重,因為“物種終結(jié),不是在最適合它的棲息地里,而是在衰退因素的最不適宜棲息地里”。因此,可以安全地假定,廣泛采用的本質(zhì)上基于對次優(yōu)棲息地內(nèi)難民物種的擔(dān)憂的保護(hù)實踐(例如烏戎庫隆國家公園內(nèi)爪哇犀的案例),在昂貴的同時,不可能產(chǎn)出期望的結(jié)果。因此,非原位活動,或者原位和非原位保護(hù)管理活動和設(shè)施的結(jié)合,例如伊比利亞猞猁和鸮鸚鵡恢復(fù),或者蘇門犀繁育中心、武廣牛(如果還有的話)和該地區(qū)的其他稀有有蹄動物的案例,是迫切需要的。
最后呼吁
印度尼西亞兩種犀牛的當(dāng)前處境悲觀地表明留存在自然棲息地中的兩個物種都只有大約12頭繁育雌性。這一嚴(yán)酷的事實強調(diào)了為確保兩個物種的生存而實施非原位繁殖的關(guān)鍵重要性。這件緊急的事呼吁積極主動政策的制定,和準(zhǔn)確的決斷。在此關(guān)鍵時刻任何對采取行動的猶豫只能被歸因于與潛在債務(wù)相關(guān)的擔(dān)心和對此事缺乏應(yīng)有的重視。
致謝
我們要感謝犀牛資源中心提供的獲取最綜合的論文合集的機會,沒有它我們對爪哇犀的研究是不可能的。我們強烈鼓勵支持涉及該物種保護(hù)的相關(guān)利益者,并且提高關(guān)于會導(dǎo)致犀牛種群災(zāi)難性下降、對犀角制品無意義的需求的毀滅性影響的意識。
特別感謝要給匿名回顧者和此原稿中所用圖片的作者。我們向?qū)ξ覀冋埱笤S可做出回應(yīng)的人表示感激,并且也感激我們無法聯(lián)系,但已經(jīng)為了他們的圖片而被給予適當(dāng)款項的作者。
附:


珍藏于西安碑林博物館的獻(xiàn)陵石犀。貞觀初年林邑國(今越南)曾向唐朝進(jìn)貢犀牛,深受李淵和李世民喜愛。李淵駕崩后,李世民命工匠雕刻石犀立于獻(xiàn)陵。可以清晰地看見石犀身上的褶皺、多邊形馬賽克狀表皮、尖的上唇和短小圓鈍的唯一犀角(進(jìn)貢的犀牛因此也被稱為圓帽犀)。結(jié)合這些特征可以確定獻(xiàn)陵石犀是爪哇犀安南亞種。

成年雄性爪哇犀身后跟隨3只豺。出自論文《Photographic record of Dholes predating on a young Banteng in southwestern Java, Indonesia》
| 版權(quán)聲明: 1.依據(jù)《服務(wù)條款》,本網(wǎng)頁發(fā)布的原創(chuàng)作品,版權(quán)歸發(fā)布者(即注冊用戶)所有;本網(wǎng)頁發(fā)布的轉(zhuǎn)載作品,由發(fā)布者按照互聯(lián)網(wǎng)精神進(jìn)行分享,遵守相關(guān)法律法規(guī),無商業(yè)獲利行為,無版權(quán)糾紛。 2.本網(wǎng)頁是第三方信息存儲空間,阿酷公司是網(wǎng)絡(luò)服務(wù)提供者,服務(wù)對象為注冊用戶。該項服務(wù)免費,阿酷公司不向注冊用戶收取任何費用。 名稱:阿酷(北京)科技發(fā)展有限公司 聯(lián)系人:李女士,QQ468780427 網(wǎng)絡(luò)地址:www.arkoo.com 3.本網(wǎng)頁參與各方的所有行為,完全遵守《信息網(wǎng)絡(luò)傳播權(quán)保護(hù)條例》。如有侵權(quán)行為,請權(quán)利人通知阿酷公司,阿酷公司將根據(jù)本條例第二十二條規(guī)定刪除侵權(quán)作品。 |